
ベランダに居座る鳩に悩まされていませんか。ネットやSNSでは様々な対策が紹介されていますが、本当に効果があるのか疑問に思うことも多いはずです。実は、鳩の行動をコントロールする最大の鍵は、彼らの持つ独自の感覚システムにあります。特に鳩の味覚や嗅覚といった化学感覚は、人間とは大きく異なる進化を遂げてきました。
この記事では、専門的な生理学の知見から鳩の味覚の正体を解き明かし、本当に効果のある科学的な防除方法から、少し意外な食文化の歴史までを徹底的に解説します。愛用のベランダを害鳥から守るための確実なアプローチを、一緒に学んでいきましょう。
この記事を読むことで理解できる内容は以下のとおりです。
- 人間と鳩における味覚の決定的な構造差
- 唐辛子が鳩に全く効かない理由と進化学的メリット
- 科学的に証明された鳥類用の忌避成分とTRPA1受容体の関係
- ベランダから鳩を完全にシャットアウトする複合的アプローチ
鳩の味覚の仕組みと驚きの真実
私たち人間が日々楽しんでいる「味わい」の感覚は、鳩にとっても同じなのでしょうか。まずは、最新の感覚生理学から明らかになった鳩の味覚構造について詳しく解説します。人間との驚くべき違いを知ることで、効果的な対策のヒントが見えてきます。
鳩の味覚と味蕾数の構造的特徴
鳥類の化学感覚システムは、視覚や聴覚の卓越した発達の影に隠れ、歴史的に「退化している」と過小評価される傾向にありました。かつての生物学研究においても、鳥類はほとんど匂いや味を感じておらず、単に目の前の餌を目視で判別してついばんでいるだけだと考えられていたのです。
しかし、近年の感覚生理学的なアプローチにより、鳥類は自らの生存戦略に最適化された独自の味覚システムを保持していることが明らかになっています。その中でも鳩(ハト目)は、人間の感覚指標とは大きく異なる、極限まで洗練されたミニマリスティックな構造を持っています。
化学物質を受容して味覚情報を脳へと伝達する、口腔内の「味蕾(みらい)」の総数において、人間が約7,902個から10,000個を保持しているのに対し、鳩が保持する味蕾はわずか37個に制限されています。この物理的な細胞数の絶対的差異は、食物に対する解像度が人間と比較して極めて低いことを生理学的に裏付けています。味わいを「楽しむ」というよりは、最低限の「生命維持のための選別」に特化した器官だと言えるでしょう。
さらに、哺乳類が舌の表面全体に高密度で味蕾を配置しているのとは異なり、鳥類における味蕾の分布は口腔内の特定の領域に偏在しています。鳥類全体の統計データによると、味蕾の約70%は上顎の口蓋部に集中し、残りの25%が下顎の底部および咽頭の深部に分布しているのです。
舌の先端にはほとんど味蕾が存在しないため、舐めて味を判断することは不可能です。このような解剖学的、生理学的配置は、鳩が歯を持たず、食物を咀嚼することなく瞬時に丸呑みするという食性と密接に適合しています。食物が口腔内に滞留する時間は極めて短く、唾液の分泌量も極めて少ないため、舌先での微細な味の判別は必要とされません。
むしろ、食物が食道へと送り込まれる嚥下の最終段階(口蓋や咽頭床)において、最低限の栄養価や致命的な有害物質を瞬時にスクリーニングするための合理的な進化学的適応の結果であると考えられます。
さまざまな生物の推定味蕾数比較
| 生物種 | 推定味蕾数(個) | 口腔構造および食性との相関 |
|---|---|---|
| 牛 | 約 25,000 | 植食性(反芻動物)。植物に含まれる複雑な化学成分や毒素を精密に選別します。 |
| 人間 | 約 7,902 〜 10,000 | 雑食性。複雑な食文化に適応した極めて高度な味覚識別能力を保持します。 |
| カラス | 約 400 | 雑食性。鳥類の中では比較的多くの味蕾を持ち、多様な対象から餌を判別します。 |
| パロット(インコ等) | 約 300 〜 500 | 果実・種子食。果実の成熟度や糖分、木の実の脂質を識別するため発達しています。 |
| ニワトリ | 約 24 〜 767 | 穀物・昆虫食。砂礫地で素早く種子や微細な餌を識別する能力を持ちます。 |
| 鳩(ハト) | 約 37 | 主に穀物(種子)食。丸呑みが主体であり、極小の味蕾を咽頭部に集約させています。 |
鳥類における五味の受容能力
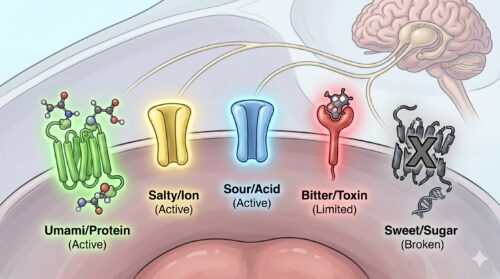
脊椎動物における基本五味(甘味、酸味、苦味、塩味、旨味)の受容能力の歴史を辿ると、鳥類は進化学的な系統樹においてドラスティックな「味覚受容体の喪失と再編成」を遂げています。そもそも、これら五つの基本味は、生物が生命を維持する上で必要不可欠な栄養源(糖、アミノ酸、ミネラル)や、体内の酸・塩基平衡を脅かす酸、そして生命の危機となる毒物(アルカロイドなど)を感知するための警報装置として、数億年をかけて獲得してきた精密なセンサー群です。
しかし鳥類は、恐竜から進化して空を飛ぶための軽量化を突き詰めるプロセス、あるいはその特異な食生活の変遷に伴って、一部の重要な受容体遺伝子を根底から失うことになりました。
鳩にとってもこの受容能力の制限は完全に適用されており、彼らは人間とは全く異なる五味の受容世界に生きています。わずか37個という驚異的に少ない味蕾の中で、彼らは「生き残るために真に必要な最低限の刺激」のみを選択的に感知し、余計な感覚をノイズとして排除しているのです。
これほど少ない感覚器官でありながら、なぜ鳩が自然界においてこれほどの繁殖力を示し、人間の生活圏を侵食するほどに適応できているのか。その謎を解き明かすためには、彼らのDNAに刻まれた味覚受容体の進化的歴史と、その独特な再編プロセスの裏側に隠された生存戦略を科学的に紐解いていく必要があります。
甘味受容体を失った進化の過程
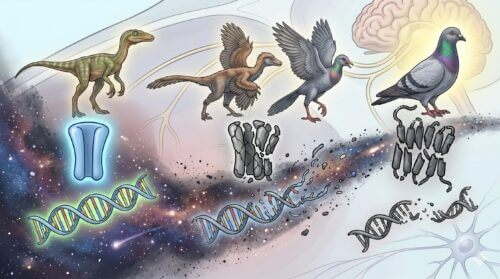
特に、炭水化物(最も効率的なエネルギー源)を感知するための甘味受容体を形成する必須遺伝子「T1R2(Tas1R2)」は、恐竜を祖先とする鳥類の進化の極めて初期段階で完全に破壊され、ゲノム中から欠損しました。したがって、現生するすべての鳥類は、遺伝子レベルで哺乳類と同等の甘味受容体を喪失しているのが大きな特徴です。甘いケーキや果実を食べても、人間のように「甘くて美味しい」と感じる脳内報酬システムは、鳥類には基本的には備わっていません。
しかしながら、野生下においてはハチドリのように花の蜜を主食とする種や、スズメ目(ソングバード)のように果実の甘味を好む種が多数存在しています。近年の最先端分子生物学的研究により、これらの特殊な食性を持つ鳥類は、本来アミノ酸(タンパク質)を認識するためのセンサーである「T1R1-T1R3(旨味受容体)」を、何千万年もの進化の過程で「リツーリング(機能転用)」し、糖類をも感知できるように受容体の結合特性を構造変化させたことが判明しました。
つまり、旨味の受容体を改変して、失われた甘味の知覚能力を二次的に「再獲得」したのです。これは平行・収斂進化の極めて稀なケースとされています。
一方で、鳩(ハト目)はこのハチドリやスズメ目が成し遂げたような「旨味受容体の甘味受容への転用」という遺伝的変異の系譜には含まれていません。鳩の口腔内においては、旨味に関連する「T1R1」および「T1R3」遺伝子はそのままの形で保存されており、本来の役割であるアミノ酸やペプチドといったタンパク質系の栄養素を検出する機能にとどまっています。このため、鳩は砂糖溶液などの甘味物質に対して積極的な知覚反応や嗜好性を示さず、エネルギーの検知を別の感覚(視覚的な色の判別や触覚)に依存しています。
さらに、有害な植物アルカロイドなどの毒物を検知するための苦味受容体「T2R」群の遺伝子保有数に関しても、鳥類間での多様性が激しいものです。野生の昆虫を主食とする種が12個もの機能的なT2R遺伝子を保持しているのに対し、鳩はわずか1個のT2R遺伝子しか持たないことが実証されています。
これは、鳩が味覚による多種多様な化学物質のスクリーニング能力を自ら大幅に制限し、その代わりに極めてシンプルな穀物や種子といった食性に自らを特化させ、必要最小限の受容能力で生理的コストを最適化していることを証明しています。
唐辛子を平気で食べる理由
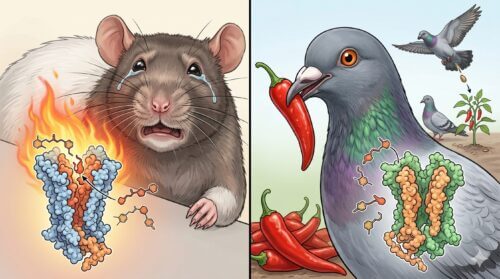
「鳩を含む鳥類は唐辛子を平気で食べる」という現象は、生物学的な共生関係および侵害受容器(痛みセンサー)の特異性を説明する上で、最も頻繁に引用される代表的な事例の一つです。人間を含む多くの哺乳類は、唐辛子を食べると強い熱さと鋭い痛み(辛味)を覚えます。
これは口内の感覚神経に発現している「TRPV1受容体」が、唐辛子の辛み成分である「カプサイシン」と特異的に結合し、これを「43℃以上の熱刺激」や「物理的な組織損傷」と同様の危険信号として脳へ伝達するためです。つまり、哺乳類にとっての辛味とは、味覚ではなく「熱痛覚」そのものなのです。
しかしながら、鳥類の保持するTRPV1受容体は、アミノ酸配列の構造的相違によりカプサイシン分子との結合部位を完全に失っています。その結果、鳥類においてはカプサイシンがTRPV1の活性化シグナルを引き起こすことはなく、どれほど高濃度の唐辛子を摂取しても口内で熱さや痛み(辛味刺激)を全く感じることがありません。鳥類にとって、唐辛子は単に栄養価の高いマイルドな果実および種子にすぎず、何の抵抗もなく平気で嚥下・消化されます。
この非感受性は、植物(トウガラシ属)と鳥類の間で何百万年にもわたって培われた精緻な進化生態学的戦略を内包しています。唐辛子は、臼歯で種子を噛み砕いて完全に死滅させてしまう哺乳類に対しては、カプサイシンという化学的警告物質を用いて自らを防御し、食害を排除します。
一方で、歯を持たずに種子を丸呑みし、飛行移動しながら消化管を通じて生存可能な種子を糞とともに遠隔地へ排泄してくれる鳥類に対しては、カプサイシンシグナルを無効化することで積極的に自らを提供し、生息域を拡大するための散布者(ディスパーサー)として活用しているのです。この絶妙な共生関係は、自然界の知恵の結晶と言えます。
実用的な害獣防除への応用
唐辛子はビタミンA前駆体であるカロテン、ビタミンC、抗酸化作用に優れたビタミンEなどが極めて豊富で、鳥類にとっては優れた健康維持源です。そのため、飼育下にあるインコやオウム、あるいは鳩のサプリメントやおやつとして唐辛子を与えることは強く推奨されています。
また、屋外の野鳥用フィーダー(餌箱)に乾燥赤唐辛子やそのフレークを意図的に混入させることにより、鳥類の健康には一切の影響を及ぼすことなく、鳥の餌を横取りするリスやネズミ、アライグマなどの哺乳類害獣だけを選択的に排除・撃退するというスマートな害獣管理(ペストコントロール)が世界中で採用されています。
※警告:加工されたホットソースやペッパースプレーなどは塩分(ナトリウム)濃度が極めて高く、鳥類が致命的な食塩中毒を起こすリスクがあるため、鳥よけや給餌の際にこれらの調味料を使用することは厳禁です。
アリルイソチオシアネート(ワサビ成分)とTRPA1受容体の活性化
とTRPA1受容体の活性化.jpg)
カプサイシンに対して生理的な不応性(反応しない性質)を示す鳥類ですが、ワサビの主成分である「アリルイソチオシアネート」のような特定の刺激物質に対しては、全く異なる過敏な反応を示します。
哺乳類において、ワサビを口にしたときのあの鼻にツーンと抜ける激しい刺激や、涙が出るような痛みは、主に「TRPA1受容体(ワサビ受容体)」を介して感知されます。このTRPA1受容体は、刺激的な化学物質だけでなく、冷感や痛覚などを総合的に感知する高感度マルチセンサーです。
鳥類のTRPA1受容体もまた、このアリルイソチオシアネートに鋭敏に反応する仕組みを保持しています。鳥類はカプサイシンによる熱や痛みを全く感じない一方で、ワサビ由来の化学刺激に対しては不快感や激しい「痛み」を覚えることが近年の研究で明らかになっています。
ワサビを混ぜた餌を鳥に与えると、激しく頭を振って吐き出したり、以降その餌を避けるような強い学習効果を伴う忌避行動が見られます。この性質は、哺乳類とは異なる独自の痛覚感知・化学物質防御システムが鳥類に存在していることを示しており、鳥類向けの特殊な感覚刺激性忌避対策を設計する上での重要な基礎知識となっています。
アントラニル酸メチルの忌避効果
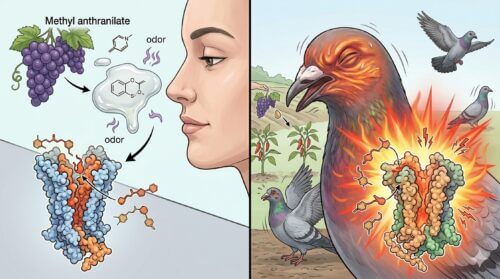
カプサイシンが効かない鳥類に対し、極めて高い化学的忌避効果を示す成分として実用化されているのが「アントラニル酸メチル(メチルアントラニレート)」です。アントラニル酸メチルは、コンキョウやブドウなどの果実や植物に自然に含まれる安全性の高いエステル化合物であり、人間にとってはブドウ風味の甘い香料や食品添加物、香水原料としても承認されている極めて身近な物質です。
しかし、この物質は鳥類に対してのみ、選択的かつ強力な不快感と苦悶を与える特殊な作用機序を持っています。海外では、農耕地や芝生(ゴルフ場)、あるいは飛行場などの滑走路周辺に散布され、深刻な農業被害や命に関わるバードストライクを防ぐための「鳥忌避剤」の主成分として広く活用されています。
アントラニル酸メチルが鳥類に対してこれほど強力な忌避を促す生理的背景には、痛覚や化学刺激を感知するセンサーである「TRPA1受容体」の極めて特殊な活性化メカニズムが関与しています。鳥類のTRPA1は、哺乳類とは異なり「高温(熱刺激)」に対しても鋭敏に反応するという、両生類や爬虫類に近い変温動物型の温度感受性を進化学的に維持しています。アントラニル酸メチルは、この鳥類特有のTRPA1受容体に特異的に配位・結合し、感覚神経に強力なチャネル開放シグナル(危険信号)を入力するのです。
この結果、アントラニル酸メチルを吸引、あるいは口にした鳥類は、実際の組織損傷がないにもかかわらず、口腔内や眼、気道に激しい「刺すような痛み」と「焼け付くような熱さ(灼熱感)」を化学的に錯覚し、驚愕して即座に対象エリアから撤退します。
このTRPA1の化学的相互作用に関するアミノ酸残基は、近年の共同研究によって分子レベルで特定されており、ニワトリやハトなどのモデルケースにおいても高確率での忌避反応(飲水拒否や摂食抑制)が証明されています。この受容体活性化をベースとして設計された商業用の「鳩よけ忌避剤」(ジェルタイプやスプレーなど)は、高分子キャリアにアントラニル酸メチルを内包して徐々に放出させることで、長期間にわたり鳩の感覚神経を強力に刺激し続け、強力な学習効果を伴う確実な駆除効果を発揮します。
鳩の味覚と嗅覚の関係性
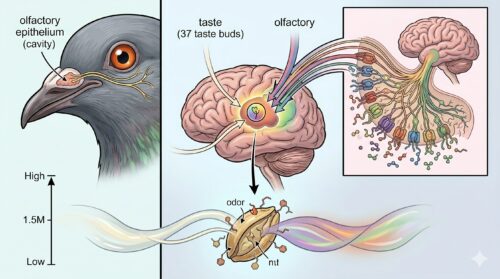
鳩の生存戦略において、極端に制限された「わずか37個の味蕾しか持たない味覚」を補っているのが、実は極めて高度に発達した「嗅覚」です。一般的に鳥類はほぼ完全に視覚と聴覚のみに依存して生活する「視覚動物」であり、化学受容器である嗅覚は進化の過程で退化しているという見解が長年にわたり定説とされていました。
しかし、現代の動物生理学および行動心理学の研究はこの仮説を根底から覆しており、鳥類、特に鳩は優れた嗅覚システムを保持し、それをフルに活用していることが証明されています。
口腔内の乏しい味覚シグナルは、鼻孔から得られる鋭敏な嗅覚シグナルと脳内で統合され、餌の選別や環境の危険性を判断する重要な「風味(フレーバー)」の情報へと昇華されています。たとえば、鳩が特定の餌を食べる際、目視による形の判別だけでなく、餌から漂うわずかな有機揮発化合物を鼻孔でスキャンし、かつ口に含んだ瞬間に咽頭部の味蕾で有害なアルカロイド(苦味)の有無を判定するという、二重のセキュリティシステムを稼働させているのです。
このように、鳩の化学感覚は味覚単体では未発達に見えますが、嗅覚と脳内で高度に統合・相互補完されることで、人間の想像を超える極めて洗練された「環境防衛および採餌ナビゲーションシステム」を形成しています。この感覚の結びつきを理解することが、ベランダ防除を成功に導くための必要不可欠なアプローチとなります。
鳩の味覚を活かした防除と食文化
鳩が持つ独自の感覚生理学を理解することは、都市環境における効果的な鳥害防除、そして伝統的な食文化における味覚設計の双方において極めて有益です。ここからは、実践的なベランダ対策や、食材としてのハト肉の科学的な特性について迫っていきます。
鳩の帰巣本能と嗅覚ナビゲーション
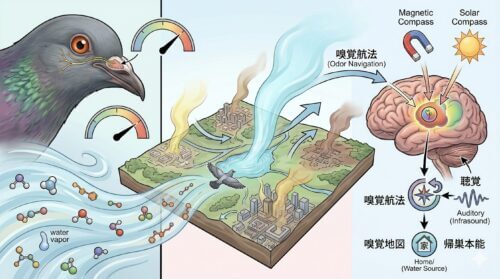
現在の動物生態学の研究は、鳥類の嗅覚退化説を完全に否定しており、特に鳩(ドバト・伝書鳩など)が極めて高度で機能的な嗅覚システムを保持し、それを自らの生命線である「帰巣本能」に直接結びつけていることを証明しています。
鳩の帰巣本能は、数万キロメートル離れた見知らぬ地点からでも、地磁気を検知する「磁気コンパス」や、偏光電磁ベクトルを検知する「太陽光偏光コンパス」、低周波を頼る「聴覚」、そして目印となる建物の記憶(視覚)をマルチモーダルに統合することで達成されます。
そしてこれらを支える強力なナビゲーション技術の一つが、広域の大気に漂う有機化学物質の微細な組成と風向から位置関係をマッピングする「嗅覚航法(嗅覚ナビゲーション)」です。鳩は左右の鼻孔で異なる大気の化学物質濃度(匂いのグラデーション)を三次元的に把握し、自らの嗅覚地図(認知地図)と照合して自身の現在座標を弾き出しているのです。
また、鳩は「水の匂い」(周辺大気の湿度や、水域に由来する微細な揮発性有機化合物)に対して極めて鋭い感受性を持っています。このため、都市部の生活空間において、鳩が公園の水場や河川、エアコンの室外機裏といった「水源」に近いエリアを巣作りの候補としてセレクトする要因となっています。
一度その場所で安全な繁殖に成功すると、鳩はそこを「自身の絶対的な縄張り」と認識します。物理的に巣を取り壊したり、あるいは綺麗に清掃を施したとしても、鳩は自らの尿腺(尾脂腺から分泌される羽油)などから発せられる個体固有の臭気シグナルや記憶された嗅覚マップを基点として、帰巣本能に突き動かされ、同一の場所へ何百回でも執念深く帰還を試みるため、表面的な対策のみでは解決が困難になります。








