日常の中でムカデに遭遇したとき、そのおぞましい姿に驚くと同時に、ふと「エビやカニなどの甲殻類と何が違うのだろう」と疑問に思ったことはありませんか。
実は学校の理科で習うように、どちらも背骨のない節足動物という大きなくくりに属しています。
しかし、ネット上でムカデと甲殻類の共通点や、ムカデと甲殻類のアレルギーに関する情報、あるいはムカデと甲殻類の仲間としてのつながりを調べてみると、直感とは大きく異なる意外な事実が次々と浮かび上がってきます。この記事では、それらの疑問を専門的な知見から分かりやすく紐解いていきます。
多足類であるムカデと、水中で暮らす甲殻類のあいだには、私たちが想像する以上に深い進化のドラマや、医学的に無視できない重篤なアレルギーの交差反応が存在しています。知っておくことで、日常生活での害虫対策や健康維持に役立つ知識を、網羅的にお届けします。
この記事を読むことで理解できる内容は以下のとおりです。
- 節足動物門におけるムカデと甲殻類の正確な系統分類と最新の研究成果
- ダンゴムシやヤスデなど混同しやすい身近な陸上節足動物の見分け方
- エビやカニのアレルギーを持つ人がムカデに対して注意すべき医学的リスク
- 古生代シルル紀から始まった節足動物の壮大な陸上進出の歴史
ムカデと甲殻類の進化と系統分類
ここでは、ムカデと甲殻類が生物学的にどのような位置づけにあり、近年の科学の発展によってその関係性がどう再定義されたのかを詳しく解説します。一見するとまったく似ていない両者ですが、遺伝子レベルで紐解くと驚くべき真実が見えてきます。
節足動物門の基本構造と分類
中学校の理科や生物の授業において、ムカデや甲殻類(エビ、カニなど)は、背骨を持たない「無セキツイ動物」の中の「節足動物門(Arthropoda)」に属するグループとして一括して導入されます。
節足動物は、キチン質からなる硬い外骨格と関節を持つ付属肢(関節肢)を共有形質とし、筋肉が外骨格の内側を支点(付着点)として強固に結合するという、私たち脊椎動物とは根本的に異なる身体構造を有しているのが大きな特徴です。
この強靭な外骨格は、内部の柔らかい臓器を保護する防具であると同時に、陸上生活においては体内の水分蒸発を防ぐための決定的な「盾」として機能しています。
この節足動物門は、現在までに記載されている現生種だけで約110万種に達し、地球上の全動物種の85%以上を占める地球上で最も繁栄に成功した巨大な分類群です。
その驚異的な多様性を整理するため、伝統的な比較形態学の分野では、体節の癒合パターン(合体節化)や肢の構造を基準に、節足動物門を大きく以下の4つの亜門に分類してきました。それぞれが生態系において独自の進化を遂げ、現代に至っています。
伝統的な節足動物門の4大分類(亜門)
- 鋏角亜門(Chelicerata): 触角を欠き、頭部に「鋏角」と呼ばれる鋏状の口器を持つ。代表例はクモ、サソリ、ダニ、カブトガニ、ウミグモなど。
- 多足亜門(Myriapoda): 1対の触角と、多数のほぼ同質な胴部体節に脚を備える。代表例はムカデ(唇脚綱)、ヤスデ(倍脚綱)、エダヒゲムシ、コムカデなど。
- 甲殻亜門(Crustacea): 2対の触角を持ち、主に水中でエラ呼吸を行う。代表例はエビ、カニ、ミジンコ、フジツボ、ダンゴムシなど。
- 六脚亜門(Hexapoda): 1対の触角を持ち、身体が頭部・胸部・腹部に明確に区分され、脚は3対。代表例は昆虫類、トビムシ類など。
このように、従来の生物学では、ムカデなどの多足類とエビやカニなどの甲殻類は、それぞれ独立した長い歴史を持つ「亜門」として並列に扱われてきました。しかし、この平穏な4大分類の枠組みは、近年のテクノロジーの進化、特に分子生物学的なアプローチによって、根底から大きく揺るがされることになります。
単肢類説を覆した分子系統学
19世紀以来、比較形態学の世界では「昆虫(六脚類)に最も近縁なグループは多足類である」という説が、きわめて自然なものとして信じられてきました。
その最大の根拠は、昆虫もムカデやヤスデも、頭部に「1対(2本)」の触角を持ち、肢の先端が枝分かれしていない「単枝型(たんしけい)」の関節肢を共有している点にありました。これらを一連のグループとして包括する系統仮説は「単肢類説(Uniramia)」と呼ばれ、教科書にも記載されるほどの不動の定説として長らく君臨してきたのです。
しかし、20世紀末から21世紀にかけて、DNAの塩基配列解析やアミノ酸配列比較、 Hox遺伝子(体節のアイデンティティを決める制御遺伝子)の発生パターン解析といった「比較ゲノム科学」が劇的な進展を遂げたことにより、この美しい単肢類説は完全に覆されることとなりました。
遺伝子レベルでの客観的な系統解析を重ねた結果、昆虫は多足類とはまったく異なる進化の系譜をたどっており、むしろ「甲殻類の内部系統から派生した極めて近縁なグループ」であることが科学的に立証されたのです。
つまり、遺伝的な距離で見ると、昆虫はムカデよりもエビやカニ、あるいはミジンコに圧倒的に近い存在であることが明らかになりました。この発見は、これまでの分類学における最大のコペルニクス的転回といえます。
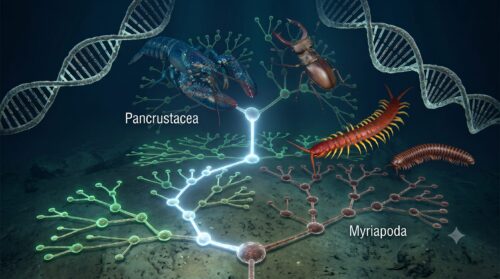
この分子系統学の発見により、従来の「甲殻亜門」は、昆虫(六脚類)を内包しない限り、共通の祖先から生まれたすべての子孫を含まない「側系統群」になってしまうことが判明しました。
そのため、現代の系統分類学では、昆虫類と甲殻類を一つに統合した強固な単系統群である「汎甲殻類(Pancrustacea)」、あるいは複眼の微細構造から「八分錘類(Tetraconata)」と呼ばれる高次分類群を確立し、これが国際的な学術標準となっています。
汎甲殻類と多足類の系統関係
昆虫と甲殻類を統合する「汎甲殻類」というグループが単系統(同一の祖先から生まれたグループ)であることを裏付ける強力な形態的証拠として、複眼の組織学的な微細構造が挙げられます。
彼らの複眼を構成する個眼の内部には、「8分割される硝子体(八分錘:はちぶんすい)」と呼ばれる光を取り込む組織が完全に共通して存在しています。
この極めて特殊な光学構造は、地球上のあらゆる動物群の中でも昆虫類と甲殻類のみが共有している固有の形質(共有派生形質)であり、両者が共通の祖先から進化した決定的な証拠となっています。
その一方で、ムカデなどの多足類は、この汎甲殻類という大グループが進化するはるか以前の、極めて早い段階で節足動物のメインストリームから分岐した独自の系統(多足亜門)です。近年の一部の分子系統解析では、多足類はむしろクモやサソリ、ダニなどの「鋏角類」に近い系統的位置にあるとする説(多足鋏角類/Paradoxopoda仮説、あるいはMyriochelata仮説)も有力視されています。
これは、外見や生息環境(陸上)からは、ムカデと昆虫が同じ多足の虫として非常に近く見えても、進化の系譜においてはムカデよりも「海のエビやカニ」の方が昆虫に圧倒的に近いという、一見すると直感に大きく反する系統関係を示しています。
生物の進化というものは、外見的な相似(相似器官)に惑わされず、ゲノムという真実の設計図を見ることで初めてその壮大なドラマが明らかになるのです。
付属肢や触角に現れる形態的差異
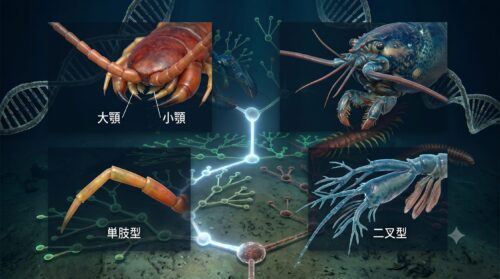
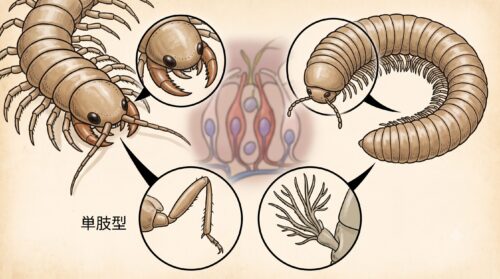
多足亜門(ムカデやヤスデ)と甲殻亜門(エビやカニ)を解剖学的に識別する上で、実用上最も重要であり、かつ進化のプロセスを如実に語ってくれる部位が「頭部の触角の対数」と「付属肢(脚)の分枝構造」です。
まず触角について比較すると、陸上で生活する昆虫類やムカデ類は、頭部に「1対(2本)」の触角しか持っていません。しかし、甲殻類は原則として「2対(4本)」の触角を備えています。
これらは「第1触角(アンテヌラ)」および「第2触角(アンテナ)」と呼ばれ、水中で化学物質(匂い)を感知したり、水の流れや振動を感じ取ったりするための高精度なセンサーとして別々に機能しています。陸生甲殻類であるダンゴムシなどでは、第1触角が肉眼で見えないほど微小に退化していますが、解剖学的には確かに2対の触角が存在しています。
さらに決定的な相違点は、付属肢の基本設計です。ムカデや昆虫の脚は、分枝を持たず一本の軸が関節で連なる「単枝型(たんしけい)」です。これは、陸上の硬い地面を効率よく歩行し、重力に抗って身体を支えるために、無駄な枝分かれを廃して強度を高めた陸上適応の結晶です。
一方、甲殻類の付属肢は、肢の基部(原節)から外側へ伸びる「外肢(がいし)」と内側へ伸びる「内肢(ないし)」に分岐する「二叉型(にさけい)」を基本構造としています。
水中で暮らす甲殻類は、この二叉構造を利用して、一方の枝を「歩行用や遊泳用のパドル」として使い、もう一方の枝に「呼吸用のエラ(鰓)」を発達させるなど、一つの脚に複数の役割を同時に持たせるという、水中ならではの合理的かつ複雑な形態変化を行っているのです。
このように、それぞれの生息環境における物理的な要求(重力、浮力、酸素濃度、媒体の密度)の違いが、単枝型と二叉型という関節肢の根本的な設計変更をもたらしました。以下の表は、各節足動物の亜門における主要な形態的特徴を網羅的に比較したものです。
| 分類群(亜門) | 触角の対数 | 付属肢の基本設計 | 口器および頭部構造 | 身体の区分 |
|---|---|---|---|---|
| 多足亜門 (ムカデ、ヤスデなど) | 1対(2本) | 単枝型(分岐なし) | 大顎1対・小顎2対 (ムカデは顎肢に毒腺を持つ) | 頭部と多数の胴部体節 (明瞭な胸腹の区別なし) |
| 甲殻亜門 (エビ、カニなど) | 2対(4本) (陸生種は退化傾向あり) | 二叉型(内外肢に分岐) (二次的に単枝化も存在) | 第1・第2触角、大顎1対 小顎2対、顎脚を持つ種も | 頭胸部(頭部と胸部が癒合) と腹部の区分が多い |
| 六脚亜門 (昆虫類など) | 1対(2本) | 単枝型(分岐なし) | 大顎1対・小顎1対 下唇1枚(小顎の融合体) | 頭部・胸部・腹部 (明確な3区分) |
| 鋏角亜門 (クモ、サソリなど) | なし(触角を欠く) | 主に単枝型 | 鋏角1対(牙や鋏に変形) 触肢1対(感覚や交尾用) | 頭胸部と腹部 (または全身が融合) |
ダンゴムシやヤスデの識別方法

一般の読者が「この目の前の多足生物はムカデなのか、それとも甲殻類なのか」という現実的な疑問を抱く現場は、大抵の場合、庭の植木鉢の裏、落ち葉の下、あるいは石をひっくり返したときといった身近な土壌環境です。そこには多種多様な「足の多い生き物」がひしめき合っており、私たちの目を惑わせます。
特に混同されやすい代表格が「ダンゴムシ(オカダンゴムシ)」や「ワラジムシ」です。平たく多足なその姿から、ムカデやヤスデの仲間、あるいは不快な昆虫の一種と捉えられがちですが、これらは生物学的には甲殻亜門・軟甲綱・等脚目(ワラジムシ目)に属する「陸生甲殻類」、すなわちエビやカニのダイレクトな親戚です。
ダンゴムシの身体を詳細に観察すると、エビの腹部にある脚(腹肢)や胸部にある脚(胸肢)に相同する、歩行のための「歩行肢」を合計で「7対(14本)」備えています(頭部にあるあご脚を含めると計8対)。
さらに、甲殻類である証拠として、顔の前には2対(4本)の触角がありますが、第1触角が肉眼では見えないほど超微小に退化しているため、外見上は1対(2本)の長い第2触角のみがピコピコと動いているように見えます。彼らは落ち葉などの有機物を食べて分解する無害な腐植食性(デトリタス食性)の生物であり、土壌を豊かにする益獣としての側面が強い存在です。
一方、ダンゴムシと並んでよく見かける「ヤスデ(倍脚綱)」は、刺激を与えると蚊取り線香のように身体を丸める種(タマヤスデなど)も存在するためダンゴムシと極めて混同されやすいですが、こちらは真の多足類です。ヤスデを識別する最大の特徴は、その脚の生え方にあります。
ヤスデの胴体は「2つの体節が進化の過程で1つに癒合した【倍体節(ばいたいせつ)】」という構造になっており、そのため基本的には1つの体節から「2対(4本)」の脚が生えています。これは、1体節から「1対(2本)」の脚しか生えていないムカデ(唇脚綱)や、7対の歩肢が均等に並ぶ等脚類(ダンゴムシ)と見分けるための決定的なポイントです。
また、注意すべき「ムカデ」は肉食のハンターであり、鋭い「毒牙(顎肢)」を頭部の直後に備えています。さらにムカデ類の脚の総対数は、奇妙なことに発育異常や事故による欠損などの例外を除き、進化のルールとして「常に奇数対(21対、23対、ゲジでは15対など)」になるという独自の発生学的・遺伝的特性を持っています。
偶数対の脚を持つ多足生物を見かけた場合、それはムカデではなく、ヤスデや等脚類などの別グループである可能性が極めて高いといえます。
ムカデエビ綱が示す進化の証拠
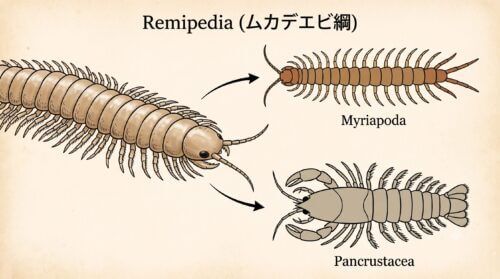
「ムカデ」と「甲殻類」という、一見すると生息環境も形態もかけ離れた両者をつなぐ、生物学史上「最も衝撃的なミッシングリンク」と評される存在が、1981年にアメリカの研究者ジル・イエガー(Jill Yager)によって発見された「ムカデエビ綱(Remipedia:レミペディア)」です。
彼らはカリブ海のバハマ諸島、カナリア諸島、西オーストラリアといった特定地域の、海水と淡水が奇跡的なバランスで分離・共存する沿岸部の海底洞窟「アンキアライン洞窟」という極めて特殊な閉鎖環境にひっそりと生息しています。ここは酸素がほとんどなく、有機物も極めて乏しい貧酸素・貧栄養環境であり、他の一般的な生物が生存できない、まるで太古の地球を思わせる限界環境です。
ムカデエビは、その名の通り、姿形が陸上の「ムカデ」に酷似しています。体長は1.5cm〜4cm程度で、頭部と、最大で42節にも及ぶ非常に細長い胴部のみで構成され、エビやカニに見られるような「胸部」と「腹部」を隔てる境界が一切ありません。
そして、すべての胴体節に、まるでボートのオールの形状をした「二叉型胴肢(泳脚)」が左右対称に美しく並んでいます。この、すべての体節がほぼ同じ形態を繰り返す「同規的体節制(どうきてきたいせつせい)」は、節足動物の祖先が数億年前に持っていたであろう原始的なデザインを色濃く残している生きた化石の特徴です。
驚くべきことに、最新の分子系統解析や発生学的アプローチにおいて、この深海の暗闇に潜むムカデエビ綱こそが、陸上で約100万種以上という爆発的な大繁栄を遂げた「六脚類(昆虫綱)」の姉妹群(最も近縁な共通祖先から分かれた血統)であることが科学的にほぼ確実視されています。
これはすなわち、「海の中でムカデエビのような姿をした祖先の一部が、長い年月をかけて陸上への適応を遂げ、今日の昆虫類へと劇的な進化を遂げた」という、生物の陸上進出のミッシングリンクを埋める極めて重要かつ動かぬ証拠なのです。
ムカデエビの神経構造や卵巣における卵形成プロセスは、昆虫への進化の過渡期を示す学術的価値がきわめて高く、現代の生物学における一線級の研究テーマとして今なお盛んに分析されています。
ムカデと甲殻類の共通アレルゲンと食味
ここからは、実生活において極めて重要な医療・健康面のリスクであるアレルギーの交差反応と、意外と知られていないムカデやゲジの食用としての特性や食味について詳しく掘り下げていきます。
トロポミオシンによる交差反応
「ムカデ 甲殻類」というキーワードで検索を行うユーザーにとって、単なる知的好奇心を超えて、時には命に関わるほどの重要性を持つのが「アレルギーの交差反応(交差抗原性)」です。
現在、成人における食物アレルギーの中で「エビ」や「カニ」を原因とする甲殻類アレルギーは極めて頻度が高く、かつ発症した場合は呼吸困難や血圧低下、意識障害を伴う重篤な「アナフィラキシーショック」に至る危険性が非常に高いことで知られています。
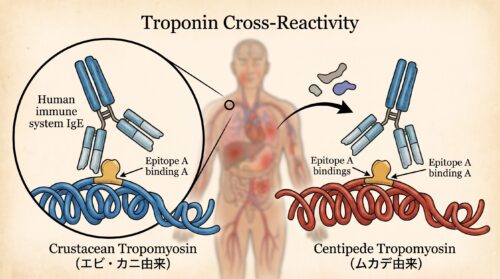
この甲殻類アレルギーを引き起こす主犯格となるアレルゲン(抗原物質)が、甲殻類の筋肉中に豊富に含まれる筋収縮制御タンパク質「トロポミオシン」です。
このトロポミオシンというタンパク質は、節足動物や軟体動物、環形動物といった無脊椎動物全般の筋肉を動かすために必須の物質であり、驚くべきことに、そのアミノ酸配列(一次構造)は進化の過程でほとんど変化せず、高度に保存されています。
つまり、エビやカニが持つトロポミオシンと、陸上の「ムカデ」や「昆虫(コオロギやイナゴなど)」、「ダニやゴキブリ」が持つトロポミオシンは、分子レベルで見ると遺伝的に酷似した構造、すなわち免疫系から見れば「ほぼ同じ形状の鍵(エピトープ)」を持っています。
そのため、エビやカニに対して過敏に反応するIgE抗体を体内に持っている人が、ムカデの体液や成分に接触・吸入・摂取した場合、体内の免疫システムがこれを「エビやカニと同じ敵が侵入してきた!」と誤認し、強力なアレルギー拒絶反応(交差反応)を爆発的に誘発してしまうのです。
この交差抗原性の問題は、単に「触らなければいい」というレベルに留まらず、住環境におけるダニやゴキブリの糞・死骸を吸い込むこと(ハウスダスト)によって無自覚のうちに感作(アレルギー準備状態)が成立し、人生で初めてエビを食べた瞬間に、いきなり重篤な甲殻類アレルギーを発症するという悲劇的なパターンを引き起こす原因にもなっています。
アレルギー症状は個人の体質によって大きく左右されるため、正確な一次情報は、専門機関のガイドライン等を参照することが不可欠です。
(参考:一般社団法人日本アレルギー学会運営・厚生労働省補助事業『アレルギーポータル』)
昆虫食や漢方薬に潜むリスク
このアレルゲンである「トロポミオシン」の恐るべき物理化学的特性として、「極めて高い耐熱性」および「強固な化学的安定性」が挙げられます。一般的な食物アレルゲン(卵や牛乳のタンパク質など)は、加熱処理を施すことで立体構造が熱変性し、アレルギー反応を起こしにくくなる特性があります。
しかし、トロポミオシンは非常に頑丈なコイル状の2本鎖構造(αヘリックス)を形成しており、熱を加えても簡単には破壊されません。それどころか、加熱調理によって一度構造がほどけても、温度が下がると再び元の強固な立体構造に自動的に再構成(リフォールディング)されるという、信じがたいタフさを持っています。
そのため、「よく火を通したから安全」「茹でたからアレルギーは起きない」という自己判断は、トロポミオシンに対してはまったく通用しません。
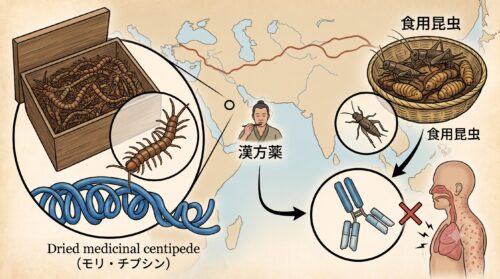
近年、持続可能なタンパク源として注目を浴びる「コオロギ食」などの昆虫食において、甲殻類アレルギーを持つ人が食べてアナフィラキシーを発症する事故が多発しており、製品パッケージには「エビ・カニ類似成分を含む」という警告が記載されるようになりました。
また、東洋医学・漢方において「蜈蚣(ごこう)」と呼ばれる乾燥ムカデを用いた生薬や、民間療法で痛み止めとして重宝されてきた、生きたムカデを強度の高いアルコールに漬け込んだ「ムカデ酒(蜈蚣酒)」、ごま油に漬けた「ムカデ油」なども極めて危険です。
トロポミオシンは、アルコール溶液中や油の中でも長期間にわたり分解されずに溶け出し、その活性を保ち続けます。甲殻類アレルギーを持つ人がこれらの薬用酒を一口でも飲んだり、皮膚の傷口にムカデ油を塗布したりすると、皮膚炎のみならず全身性のアナフィラキシーショックを誘発する引き金になります。
民間療法を試みる前には、必ず専門医への相談を徹底してください。
トロポミオシンアレルギーの重要チェックポイント
- 加熱調理による無毒化は不可能: 熱変性に対する極めて強い耐性を持ち、加熱してもアレルゲン性は低下しません。
- アルコールや油でも失活しない: ムカデ酒やムカデ油といった抽出液の中にも、活性を持ったまま永続的に溶け込みます。
- 乾燥粉末による吸入リスク: ムカデや昆虫、ダニの死骸が乾燥して粉末状になり、空気中を漂うものを吸い込むだけでも、気管支喘息やアレルギー性鼻炎、ひいては食物アレルギーとしての甲殻類過敏症を誘発・悪化させる原因になります。
咬傷によるアナフィラキシー
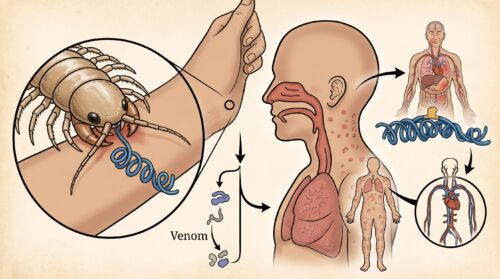
ムカデに関する医学的リスクの中で、臨床の現場で極めて重大視されているのが、ムカデに「咬まれる(咬傷)」ことによって発生する急性アレルギー反応です。
日本国内には、最大15cm近くに達するトビズムカデやアオズムカデといった大型種が生息しており、これらは初夏から秋にかけて活発に活動し、しばしば民家に侵入して寝具や衣服の中に潜り込み、人間を激しく咬みます。
通常のムカデ咬傷であっても、その顎肢(がくし:毒牙)から注入される毒素(ヒスタミン、セロトニン、ヒアルロニダーゼ、溶血性タンパク質、神経毒ペプチドなど)の作用によって、激しい灼熱痛や発赤、局所の著しい腫れ、時にはリンパ管炎を引き起こします。
しかし、甲殻類アレルギーを罹患している人の場合、これら通常の局所毒性反応とは比較にならない、命に関わる「全身性アナフィラキシー」が引き起こされる臨床実例が数多く報告されています。
これは、ムカデに咬まれた傷口から、注入された毒成分や、ムカデの体液・口器周辺に付着しているトロポミオシンおよび類似のアレルゲンタンパク質がダイレクトに毛細血管を通じて体循環へと侵入するためです。
すでにエビやカニによって体内に形成されていた「抗トロポミオシンIgE抗体」が、侵入したムカデ成分と瞬時に結合し、全身の肥満細胞から化学伝達物質(ヒスタミン等)が一斉に放出されます。
その結果、咬まれてからわずか数分〜数十分の間に、全身の蕁麻疹、喉の粘膜の劇烈な浮腫による気道閉塞(窒息リスク)、喘鳴、血圧が急激に低下するアナフィラキシーショックが誘発されます。
甲殻類アレルギーをお持ちの方は、家庭内や野外でのムカデ遭遇および咬傷被害に対して、一般の方以上に厳重な警戒を払い、万が一咬まれた場合は一刻も早く医療機関への救急受診や、所持している場合は自己注射薬(エピペン)の投与といった迅速な処置が必要です。
食用多足類の味覚と食味の対比
ムカデやゲジといった多足類は、その奇怪で不気味な見た目や、咬まれると激痛を伴う危険性から、現代社会においては激しく嫌悪される「不快害虫」の筆頭ですが、アジアをはじめとする世界各地における「食用」としての歴史、また実際にこれらを食した探求者や研究報告を分析すると、驚くべき味覚特性が明らかになります。それは、彼らの系統分類上の遠い親戚である「甲殻類(エビやカニ)」に極めて酷似した風味を持っているという事実です。
例えば、新鮮なオオムカデを油でカラリと揚げた「ムカデの素揚げ」は、噛みしめた瞬間の外殻の食感において、エビやサワガニを丸揚げにしたときのような香ばしくパリパリとした非常に小気味よいキチン質の軽快さを楽しむことができます。
さらに噛み進めると、口の中にエビやシャコを想起させる濃厚で芳醇なアミノ酸の「旨味」がじわりと広がります。
しかし、ムカデ独自の決定的な特徴は、その後に襲ってくる強烈な「苦味」にあります。この独特の風味は、エビのような純粋な甘みとは異なり、海産物の「さんまのワタ(苦味のある内臓)」や「うなぎの肝焼き」のような、奥深いコクと薬用的なほろ苦さを併せ持った「大人の味わい」に非常に近いと評されます。
これはムカデが肉食の捕食者であり、体内に様々な生理活性物質(毒素や代謝産物)を内包していることに関連していると考えられます。
一方、より嫌われがちな「ゲジ(ゲジゲジ)」についても、実は驚くほど高い食用価値が報告されています。ゲジはムカデに比べて外皮が非常に薄く、水分含有量も適度なため、素揚げにすると、ムカデ特有の生臭さや漢方薬のような薬臭さが一切なく、完全にクセのない「極上のパリパリ・サクサク食感スナック」へと変貌します。
際立って長いその脚についても、「見方を変えれば桜エビや川エビの唐揚げの髭や脚の密集度と変わらない」と指摘され、ビールのおつまみ等として非の打ち所がない美味しさを持つことが確認されています。
これら多足類の味覚が甲殻類に非常に近いという事実は、彼らが共通の祖先から受け継いだキチン質の外骨格、筋肉アミノ酸、そしてアレルゲンでもあるトロポミオシンなどの化学的構成成分の共通性を、私たちの舌がダイレクトに証明している非常に面白い事例といえます。
生態系におけるムカデと甲殻類の「捕食・被食」関係
ムカデと甲殻類のつながりは、進化や味の共通性だけではありません。自然界の生態系、特に渓流沿いや湿潤な森林土壌においては、両者は「捕食・被食」という緊迫した関係で交錯しています。
大型化するオオムカデ類は、驚異的なパワーと毒牙を用いて、地上を這い回る昆虫のみならず、陸生甲殻類であるオカダンゴムシ、さらには水辺に生息するサワガニなどの小型甲殻類をも容赦なく襲い、その頑丈な大顎と小顎を使ってバリバリと咀嚼し、捕食します。
陸上の絶対的肉食ハンターであるムカデにとって、水辺の甲殻類は、栄養豊富な絶好の獲物(ごちそう)でもあるのです。
古生代シルル紀からの陸上適応
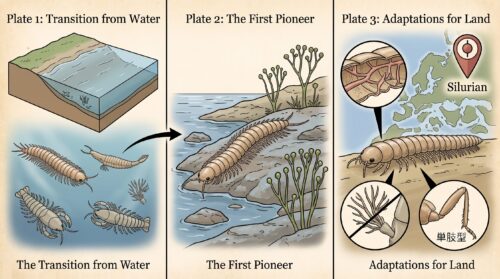
地球という生命の舞台において、「海から陸への進出(上陸作戦)」を、脊椎動物(両生類の祖先)よりもはるかに先駆けて、最初に成功させた偉大な開拓者こそが、ムカデなどの多足類を含む節足動物でした。
今から約4億4,300万年前〜4億1,900万年前の古生代シルル紀、地球の大気中には光合成を行うシアノバクテリアや藻類の働きによって酸素が十分に蓄積され、有害な宇宙線を遮断する「オゾン層」が形成されました。これにより、地表は生命が生存可能な安全な空間へと生まれ変わり、まずクックソニアやリニアといった原始的な陸上植物が湿潤な湿地帯へと進出、緑のカーペットを形成し始めました。
この湿った植物遺骸や菌類という「エサ」を追うようにして、およそ4億1,800万年前(シルル紀後期)、動物界のトップバッターとして陸上に這い上がったのが、ムカデやヤスデの祖先である最初期の多足類や、サソリの祖先(鋏角類)でした。
彼らは、海中で培った「キチン質の硬い外骨格」という、乾燥と重力から身体を守るための完璧なスーツをあらかじめ身につけていたため、陸上という過酷な新天地に最も早く適応することができたのです。
その後、デボン紀前期(約4億年前)には、汎甲殻類の共通祖先から水中で分岐した「昆虫の祖先(六脚類)」が陸上へと適応し、気管呼吸システムを発達させ、ついには空を飛ぶ「翅(はね)」を獲得して爆発的な多様化を遂げました。
この時、昆虫は多足的な多くの脚を持つ祖先から、脚の数を「3対(6本)」へと機能的に退化・特化させていったプロセスが、デボノヘキサポドゥスなどの化石記録から判明しています。
一方、甲殻類そのもののグループも、歴史上、何度も海から陸上への移行を独立して試みてきました。その中で、現代の陸上において完璧な適応と驚異的な多様化を成し遂げたのが、ダンゴムシやワラジムシが属する等脚類(陸生等脚類)です。
彼らは多足類の上陸よりもはるかに後世の時代になってから独自に上陸を果たしましたが、水中でエラ呼吸を行っていた「腹肢(ふくし)」と呼ばれる腹部の平たい付属肢を、空気中の酸素を直接取り込める微細な管状構造である「偽気管(ぎきかん:または偽肺)」へと劇的に発生変化させることで、乾燥する陸上でも生命活動を維持する呼吸システムを確立しました。
このように、最古の陸上開拓者であるムカデ、海中で甲殻類から分岐した昆虫、そして独自にエラをリフォームして上陸したダンゴムシに至るまで、節足動物の進化の系譜は、海と陸を幾度も行き来するダイナミックな歴史そのものなのです。
ムカデや甲殻類のまとめ

これまで解説してきたように、ムカデという「地球最古の陸上開拓者」の系譜から、水中での甲殻類の進化、そしてそこから分岐した昆虫類の爆発的な陸上適応、さらにはダンゴムシのように独自に上陸を果たした「陸生等脚類」のストーリーに至るまで、節足動物の進化の歩みは、海と陸という2つの巨大な世界を何億年にもわたって繋ぎ続ける、気が遠くなるほど壮大でドラマチックなものです。
私たちが日常の中で、一見全く関係がない、あるいは正反対の形状をしていると感じる「ムカデ」と「甲殻類」のあいだには、分子系統学がもたらした「汎甲殻類(昆虫と甲殻類の統合)」という驚くべき遺伝的真実が存在しています。
そして何より、医学的な観点においては、筋肉タンパク質「トロポミオシン」を共通のアレルゲンとすることによる重篤な「アレルギーの交差反応」という、私たちの健康や命に直結する非常にシビアな問題が背後で結実しています。
これらアレルギーの機序や個別のアナフィラキシーリスク、あるいは身の回りの安全対策に関する具体的な判断については、アレルギーを持つ方の健康状態や体質によって極めて多様に異なるため、決して安易な自己判断を行わず、確実な治療や正確な情報は公式サイトをご確認ください。
そして、万が一の症状発生時や、予防的な生活設計における最終的な判断は専門家にご相談ください。身近に潜む危険なハンターであるムカデの生態や進化、そして医学的なリスクを正しく理解し、毎日の安全で快適な暮らしにしっかりと役立てていきましょう。








